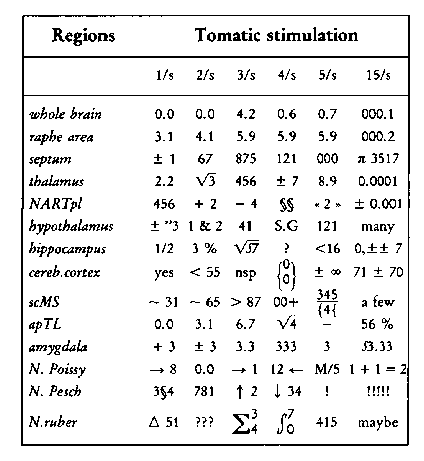
TABLE ONE. Differential responding of tomatic stimulation in the brain at different frequencies.
L'auteur étude les fois que le lancement de la tomate il provoquit la réaction yellante chez la Chantatrice et demonstre que divers plusieures aires de la cervelle elles etait implicatées dans le response, en particular, le trajet légumier, les nuclei thalameux et le fiçure musicien de l'hémisphère nord.
As observed at the turn of the century by Marks & Spencer (1899), who first named the “yelling reaction” (YR), the striking effects of tomato throwing on Sopranoes have been extensively described. Although numerous behavioral (Zeeg & Puss, 1931; Roux & Combaluzier, 1932; Sinon et al., 1948), pathological (Hun & Deu, 1960), comparative (Karybb & Szyla, 1973) and follow-up (Else & Vire, 1974) studies have permitted a valuable description of these typical responses, neuroanatomical, as well as neurophysiological data, are, in spite of their number, surprisingly confusing. In their henceforth late twenties' classical demonstrations, Chou & Lai (1927 a, b, c, 1928 a, b, 1929 a, 1930) have ruled out the hypothesis of a pure facio-facial nociceptive reflex that has been advanced for many years by a number of authors (Mace & Doyne, 1912; Payre & Tairnelle, 1916; Sornette & Billevayzé, 1925). Since that time, numerous observations have been made that have tried to decipher the tangling puzzle as well as the puzzling tangle of the afferent and/or efferent sides of the YR and led to the rather chaotic involvement of numberless structures and paths: trigeminal (Loewenstein et al., 1930), bitrigeminal (Von Aitick, 1940), quadritrigeminal (Van der Deder, 1950), supra-, infra-, and inter-trigeminal (Mason & Ragoun, 1960) afferents have been likely pointed out as well as macular (Zakouski, 1954), saccular (Bortsch, 1955), utricular (Malosol, 1956), ventricular (Tarama, 1957), monocular (Zubrowska, 1958), binocular (Chachlik, 1959-1960), triocular (Strogonoff, 1960), auditive (Balalaika, 1515) and digestive (Alka-Seltzer, 1815) inputs. Spinothalamic (Attou & Ratathou, 1974), rubrospinal (Maotz & Toung, 1973), nigro-suiatal (Szentagothai, 1972), reticular (Pompeiano et al., 1971), hypothalamic (Hubel & Wiesel, 1970), mesolimbic (Kuffler, 1969) and cerebellar (High & Low, 1968) pathways have been vainly searched out for a tentative explanation of the YR organization and almost every part of the somesthesic (Pericoloso & Sporgersi, 1973), motor (Ford, 1930), commissural (Gordon & Bogen, 1974) and associative (Einstein et al., 1974) cortices have been found responsible for the progressive building-up of the response although, up-to-now, no decisive demonstration of both the input and output of the YR programming has been convincely advanced.
Recent observations by Unsofort & Tchetera pointing out that “the more you throw tomatoes on Sopranoes, the more they yell” and comparative studies dealing with the gasp-reaction (Otis & Pifre, 1964), hiccup (Carpentier & Fialip, 1964), cat purring (Remmers & Gautier, 1972), HM reflex (Vincent et al., 1976), ventriloquy (McCulloch et al., 1964), shriek, scream, shrill and other hysterical reactions (Sturm & Drang, 1973) provoked by tomato as well as cabbages, apples, cream tarts, shoes, buts and anvil throwing (Harvar & Mercy, 1973) have led to the steady assumption of a positive feedback organization of the YR based upon a semilinear quadristable multi-switching interdigitation of neuronal sub-networks functioning en desordre (Beulott et al., 1974). Although this hypothesis seems rather seductive, it lacks anatomical and physiological foundations and we therefore decide to explore systematically the internal incremental or decremental organization of the YR, allowing a tentative anatomic model.
Control experiments were made with other projectiles, as apple cores, cabbage runts, hats, roses, pumpkins, bullets, and ketchup (Heinz, 1952).
TABLE ONE. Differential responding of tomatic stimulation in the brain at different frequencies.
Examples of responses obtained from these structures can be seen on figure 1 where temporal analysis of the spike distribution based on their Responsive-Area-Temporal-Programming (RATP) properties allowed to distinguish 3 unit subtypes: 1) units responding before the stimulation; 2) units responding during the stimulation and 3) units responding after the stimulation.
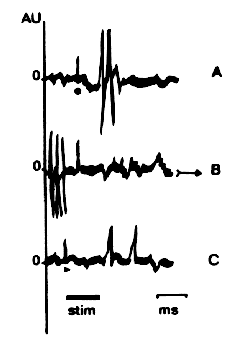
Fig. 1: Unit activity in structures responding to tomatic stimulation. Bar
indicates stimulus onset & cessation.
Calibration: 3.1416 ms. Each trace is made of the superimposition of
33.57 successive recordings.
Note the point in A, the arrow in B and the black triangle in C.
Cross-examination of responses driven by other projectiles and Ketchup stimulation are shown on figure 2 and argue unquestionably in favor of a tomatotopic organization of the YR along, between and across the NARTpl, apTL and scMS. Temporal relationships of those responses, as examplified in fig. 3, showed that the hypothesis of a clustering interdigitation of neuronal subnets is highly probable, although no experimental evidence can be given due to the relative difficulty of entering those damned structures without destroying a lot of things (Timeo et al., 1971).
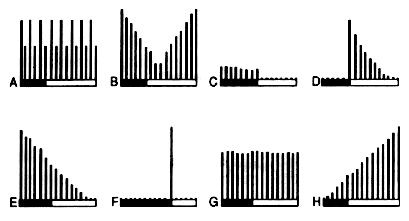
Fig. 2: Examples of response in the apTL provoked by tomato and other
throwings. Explanations in text.
A = tomato; B = apple; C = cabbage; D = hats; E = roses;
F = ketchup (kindly provided by Laroche-Ciba, Inc.); G = pumpkin; H = bullet.
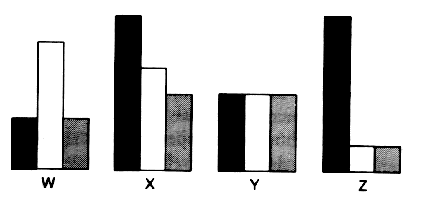
Fig. 3: Temporal relationship of the responses recorded in the YR
area.
Abscissae: arbitrary units; ordinates: international
units. Explanations in text.
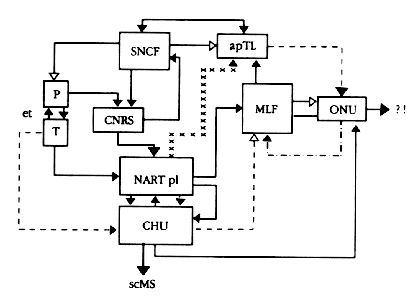
Fig. 4: Tentative anatomical model of the YR organization.
Explanations in text or elsewhere.
Black lines = inhibitory;
borken lines = interrogatory;
dashed lines = redhibitory;
stellate lines = whig-and-tory.
This work was supported by grants from the Syndicat régional des Producteurs de Fruits & Légumes, the Association française des Amateurs d'Art Lyrique (AFAAL) and the Fédération internationale des Dactylo-Bibliographes (FIDB).
The author gratefully acknowledges the helpful criticisms as well as the skilful assistance of J. Chandelier, M. De Miroschedji and H. Gautier.
Attou, J. & Ratathou, F. Laminar configuration of the thalamo-tomatic relay nuclei. Experimental study with Fink-Heimer-Gygax methods. In: The Hyperthalamus, ed. by V. Cointreau and M. Brizard, Cambridge, Oxford U.P., pp. 32-88, 1974.